Släktskapsselektion
Från Rilpedia

Släktskapsselektion är en teori som innebär att organismer uppvisar strategier som ökar den reproduktiva framgången hos sina släktingar till kostnad av sin egen reproduktiva framgång eller överlevnad. Förutsättningar för att detta ska evolvera är (a) att den reproduktiva fördelen för den mottagande individen är större än nackdelen det innebär för aktören och (b) att den altruistiska genen även finns i den mottagande individens genom.
Begreppet släktskapsselektion myntades först av John Maynard Smith 1964 [1], men teorin grundar sig på begreppet "inkluderande fitness" (eng. inclusive fitness) som William Donald Hamilton beskrev tidigare samma år. Modellen innebär förenklat att en individs gener förs vidare till nästa generation, dels genom dess egen reproduktion och dels genom ett altruistiskt beteende som resulterar i reproduktiv framgång hos individer med samma genotyp [2]. Några exempel på evolution av släktskapsselektion är varningsläten, osmaklighet hos larver, och fåglar som stannar hos sina föräldrar första året och hjälper dem med nästa kull. Nedan tas två exempel upp mer i detalj: släktskapsselektion hos sociala insekter och hos människan.
Innehåll |
Hamiltons regel
För att förklara hur ärftligt altruistiskt beteende kan uppstå utvecklade W. D. Hamilton [3][4][5] en teori som senare kom att betecknas Hamiltons regel:
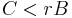
C = (eng. cost) den reproduktiva kostnaden för den agerande (altruistiska) individen.
r = (eng. relatedness) genetiskt släktskap mellan aktören (altruisten) och mottagaren.
B = (eng. benefit) den tillkommande reproduktiva fördelen som mottagaren får.
Den reproduktiva kostnaden (C) för den altruistiska individen mäts i reducerad fitness, medan den reproduktiva fördelen (B) för den mottagande individen mäts i ökad fitness. Originaldefinitionen för släktskap (r) i Hamiltons regel betecknades som ”sannolikheten att i ett slumpvist utvalt locus finna alleler som är identiska, beroende på släktskap” [3]. Genetiskt släktskap (r) mellan aktören och mottagaren kan variera från 0 (helt obesläktade) till 1 (genetiskt identiska). Således är det genetiska släktskapet mellan förälder och avkomma r = 0.5 (även i fallet mellan två helsyskon), medan r = 0.125 mellan kusiner.
Om effekten av den altruistiska individens gärningar blir att den ökar frekvensen av sina egna gener i form av en släktings avkomma, mer än vad den förlorar på att inte reproducera sig själv, kan altruistisk släktskapsselektion uppstå. Altruistiskt beteende kan också uppstå mellan grannar oavsett släktskap (som t.ex. varningsrop hos fåglar), om risken eller nackdelen den agerande individen utsätts för är väldigt liten, eller om grannarna i den närmsta omgivningen inte är alltför avlägset besläktade [3].
Mekanismer
Enligt Hamilton [3] kan altruistiskt beteende endast vara lönsamt om det är riktat mot (reproduktiva) släktingar, som har större chans att bära på den gen som kodar för det altruistiska beteendet, och om fördelen för mottagaren överväger nackdelen för aktören. För syskon måste denna fördel vara dubbelt så stor som nackdelen för att altruism ska selekteras för. Ett simplifierat exempel för detta skulle vara att en individ skulle offra sig själv för två syskon, men inte för färre. Hamilton beskrev två mekanismer på hur detta kan ske:
- För att släktskapet mellan individer ska vara högt nog för att främja altruism måste individerna på något sätt kunna känna igen släktingar och anpassa sitt beteende enligt detta, s.k. "släktskapsigenkänning" (eng. kin discrimination). Denna mekanism skulle dock vara begränsad till framför allt djur som har sexuell förökning.
- Men det kan också finnas icke-diskriminerande altruism som existerar bland organismer som inte sprider sig märkvärt utan lever tätt samman, t.ex. hos mikroorganismer som slemsvampar. Detta eftersom sannolikheten att de närmast levande individerna är besläktade är tillräckligt stor för att altruism ska vara lönsamt [6].
Sociala insekter
En grupp av organismer där man starkt kan påvisa släktskapsselektion är sociala insekter. Det beror bl.a. på att de uppvisar en högre grad av social integration än vad t.ex. sociala vertebrater gör, och att arbetarna hos denna grupp ofta är sterila vilket motsäger att individuell selektion skulle vara den enda verkande. Teorin om släktskapsselektion förutsätter att medlemmar i kolonin måste vara besläktade och att sterilitet uttrycks, och båda dessa förutsättningar finns det starkt stöd för. Ett exempel på en utmärkande form av altruism hos sociala insekter är arbetarnas aggressivitet mot inkräktare som t.ex. kan yttra sig genom att de sticks. Benägenheten att stickas är nära korrelerat till arbetarnas sterilitet, och drottningar visar inte denna typ av aggressivitet jämfört med arbetarna [7].
En stor grupp inom sociala insekter är Hymenoptera som innefattar bin, myror och getingar. Något som kännetecknar denna grupp är att de är haplodiploida, vilket innebär att det utvecklas diploida honor från befruktade ägg och haploida hannar från obefruktade ägg. Det här innebär att könskvoten kommer att vara tre reproduktiva honor på en reproduktiv hanne och att systrar är besläktade till ¾ och bröder till ¼. Honor tjänar alltså mer på att satsa på sina systrar än på sin egen avkomma, där släktskapet är ½. Drottningen i kolonin kommer fortfarande att selekteras för en 1:1 kvot eftersom hon är lika mycket besläktad med sina söner som med sina döttrar. Hannarna kommer även att ha en tre gånger så stor reproduktiv fördel än honorna i och med att könskvoten är 1:3. Denna kvot kommer därför att befinna sig i jämvikt. Trots att hypotesen om haplodiploidi verkar mycket lovande så finns det en del oklarheter. T.ex. är släktskapet mellan systrar ofta lägre än ¾ eftersom många kolonier innehar mer än en drottning, som inte är besläktade [7] .
Mänsklig släktskapsselektion
Mänskliga samhällen avviker stort från andra djursamhällen då de baseras på storkaligt samarbete mellan genetiskt obesläktade individer. Tydliga exempel på detta kan hittas i moderna samhällen (organisationer, institutioner), men även i jägar-samlarsamhällen (kooperativ jakt, kollektivt matdelande och krigsförande). Hos de flesta andra djur begränsas kooperation till små grupper, undantaget sociala insekter och nakenråttor (där släktskapet inom gruppen dock är betydande).
Mänsklig altruism skiljer sig också genom att den inte bara grundar sig på reciprok altruism och ryktesbaserad kooperation, utan även är starkt reciprok. Starka reciprokatorer bär kostanden själva för att premiera någon annan eller bli bestraffade, trots att de inte personligen gynnas av detta, varken direkt eller långsiktigt [8].
Men även hos människor har man funnit ett mer altruistiskt agerande mot närbesläktade individer, både i undersökningar där konsekvenserna av beteendet varit hypotetiska och där de varit verkliga. Hamiltons regel har kunnat appliceras på mänskligt beteende och föreslagits vara en baslinje för mänskligt altruistiskt beteende, där kultur, etik och moral, sexuell attraktion m.m. tillkommer [9].
Se även
Referenser
- ↑ Smith, J. M. (1964). "Group Selection and Kin Selection". Nature 201 (4924): 1145–1147. DOI:10.1038/2011145a0.
- ↑ Stearns, S. C.; Hoekstra R. F.: Evolution: an introduction, Oxford University Press, 2005, 2.
- ↑ 3,0 3,1 3,2 3,3 Hamilton W. D. (1963). "The Evolution of altruistic behavior". American Naturalist 97: 354–356.
- ↑ Hamilton W. D. (1964). "The Genetical Evolution of Social Behavior. I". Journal of Theoretical Biology 7 (1): 1–16. DOI:10.1016/0022-5193(64)90038-4..
- ↑ Hamilton W. D. (1964). "The Genetical Evolution of Social Behavior. II". Journal of Theoretical Biology 7 (1): 17–52. DOI:10.1016/0022-5193(64)90039-6..
- ↑ West, S. A.; Gardener, A.; Gryffin, A. S. (2006). "Quick guide: Altruism". Current Biology 16: R482–R483.
- ↑ 7,0 7,1 Queller, D. C. & Strassman, J. E. (1998). "Kin Selection and Social insects". BioScience 48: 165–175.
- ↑ Fehr, E. & Fischbacher, U. (2003). "The Nature of Human Altruism". Nature 425: 785–791.
- ↑ Madsen, E. A.; Tunney, R. J.; Fieldman, G.; Plotkin, H. C.; Dunnbar, R. Y. M.; Richardson, J. M.; McFarland, D. (2007). "Kinship and Altruism: a Cross-Cultural Experimental Study". British Journal of Psycology 98 (2): 339–359.